오스트랄로피테쿠스 아프리카누스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
오스트랄로피테쿠스 아프리카누스는 약 330만 년에서 210만 년 전 사이에 존재했던 멸종된 호미닌 종으로, 남아프리카에서 발견되었다. 1924년 레이몬드 다트가 타웅 아이를 발견하면서 처음으로 알려졌으며, 이후 미세스 플레스, 리틀 풋 등 다양한 화석이 발견되었다. 이들은 두 발로 걷는 직립 보행을 했으며, 뇌 용량은 유인원 수준이었지만, 인간에 가까운 두개골 특징을 보였다. 잡식성이었으며, 맹금류나 대형 육식 동물에게 사냥당했을 가능성이 있다.
더 읽어볼만한 페이지
- 1925년 기재된 화석 분류군 - 오스트랄로피테쿠스
오스트랄로피테쿠스는 420만 년 전부터 200만 년 전까지 아프리카에 살았던 초기 인류 속으로, 작은 뇌와 이족보행 능력을 지녔으며, 다양한 종과 화석 증거를 통해 호모 속의 조상일 가능성이 제기되는 등 인류 진화 연구에 중요한 자료이다. - 1925년 기재된 화석 분류군 - 아르히노케라톱스
아르히노케라톱스는 백악기 후기 북아메리카에 살았던 각룡류 공룡의 한 속으로, 1923년 캐나다에서 두개골 화석이 발견되어 1925년에 아르히노케라톱스 브라키옵스라는 모식종으로 명명되었으며, 속명은 '코 뿔이 없는 얼굴'을 의미하나 이는 초기 연구자의 오해에서 비롯되었다. - 오스트랄로피테쿠스 - 타웅 아이
타웅 아이는 레이먼드 다트가 발견한 약 3~4세의 *오스트랄로피테쿠스 아프리카누스* 두개골 화석으로, 초기에는 논쟁이 있었으나 후속 연구를 통해 인류 진화 연구에 중요한 초기 호미닌으로 인정받았으며, 두개골 형태와 대후두공 위치는 직립 보행 가능성을 제시한다. - 오스트랄로피테쿠스 - 오스트랄로피테쿠스 가르히
약 250만 년 전 에티오피아에서 발견된 오스트랄로피테쿠스 가르히는 450cc 정도의 뇌 용적과 큰 어금니, 소구치를 가지며, 초기 도구 사용 흔적과 함께 발견되어 도구 사용과의 연관성이 제시되고, 종명은 아파르어로 "놀라움"을 뜻한다. - 플라이오세 아프리카의 포유류 - 호모 에렉투스
호모 에렉투스는 약 200만 년 전부터 10만 년 전까지 생존하며 직립 보행을 하고 아슐리안 석기를 제작했으며 불을 사용한 최초의 인류로 추정되는 종이다. - 플라이오세 아프리카의 포유류 - 오스트랄로피테쿠스
오스트랄로피테쿠스는 420만 년 전부터 200만 년 전까지 아프리카에 살았던 초기 인류 속으로, 작은 뇌와 이족보행 능력을 지녔으며, 다양한 종과 화석 증거를 통해 호모 속의 조상일 가능성이 제기되는 등 인류 진화 연구에 중요한 자료이다.
| 오스트랄로피테쿠스 아프리카누스 - [생물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 학명 | Australopithecus africanus |
| 명명자 | 레이먼드 다트, 1925년 |
| 화석 범위 | 후기 플리오세 - 초기 플라이스토세, 330만년 전 ~ 210만년 전 |
| 동의어 | Australopithecus transvaalensis 로버트 브룸, 1936년 Plesianthropus transvaalensis 로버트 브룸, 1938년 Australopithecus prometheus? 다트, 1949년 |
| 속 | 오스트랄로피테쿠스속 |
| 특징 | |
| 두뇌 용량 | 420–510 cc |
| 키 | 약 140 cm |
| 몸무게 | 약 40 kg |
| 여성 키 | 약 125 cm |
| 여성 몸무게 | 약 30 kg |
2. 발견 및 연구사
1924년 레이몬드 다트가 타웅 아이 화석을 발견하고 '오스트랄로피테쿠스 아프리카누스'라는 학명을 부여하면서, 인류 진화 연구에 중요한 전환점이 마련되었다. 찰스 다윈은 인류가 아프리카에서 진화했을 것이라고 주장했는데, 이는 오스트랄로피테쿠스 아프리카누스 발견으로 뒷받침되었다. 그러나 당시 학계는 다트의 주장을 즉각 받아들이지 않았다.
1938년 로버트 브룸은 스테르크폰테인에서 발견된 성인 두개골 화석(Sts 5)을 '플레시안트로푸스 트란스발렌시스'로 분류하며 다트의 주장을 지지했다. 1947년 브룸과 존 T. 로빈슨은 미세스 플레스라고 불리는 화석(Sts 5)을 발견했는데, 이 화석은 유인원의 특징이 아닌 인류와 가까운 특징을 보여, 후에 오스트랄로피테쿠스 아프리카누스로 재분류되었다. 제2차 세계 대전 이후 남아프리카 여러 지역에서 추가 화석이 발견되고, 1953년 필트다운 인 화석 조작 사건이 밝혀지면서 오스트랄로피테쿠스 아프리카누스가 인류의 조상이라는 주장은 더욱 힘을 얻게 되었다.
1994년 남아프리카 스테르크폰테인 동굴에서 "리틀 풋"(StW 573)이라고 불리는 거의 완전한 오스트랄로피테쿠스 아프리카누스 골격 화석이 발견되었다. 리틀 풋의 발견은 오스트랄로피테쿠스 아프리카누스 내 종 분화 논쟁을 다시 불러일으켰다.
2. 1. 타웅 아이의 발견과 다트의 주장
1924년, 호주 출신의 해부학자 레이몬드 다트는 남아프리카 타웅의 석회암 채석장에서 발파공 M.G. 드 브루인이 발견한 어린 개체의 두개골 화석을 받았다. 이 화석은 '타웅 아이'라고 불리며, 뇌는 작았지만 대후두공의 위치로 보아 직립 보행을 했다는 특징을 보였다.[8] 다트는 이 화석이 유인원과 인간 사이의 과도기적 화석이라고 판단하여, '오스트랄로피테쿠스 아프리카누스'(남아프리카의 유인원)라는 새로운 학명을 부여했다.[8]찰스 다윈은 1871년 저서 ''인간의 유래''에서 인류가 아프리카에서 진화했을 것이라고 가정했는데, ''A. africanus''는 이러한 주장을 뒷받침하는 최초의 증거였다. 그러나 당시 학계에서는 큰 뇌 크기와 인간과 같은 특징이 비교적 일찍 발달했고, 이족 보행보다 뇌 크기가 먼저 진화했다는 생각이 지배적이었다. 따라서 다트의 주장은 즉각적으로 받아들여지지 않았다. 특히, 영국의 해부학자 아서 키스는 타웅 아이를 유인원의 어린 개체로 간주하며 다트의 주장을 반박했다.[8]
1912년 찰스 도슨이 조작한 필트다운 인 화석 사건과 중국에서 발견된 베이징 인(''Homo erectus pekinensis'') 화석은 인류의 기원이 아프리카가 아닌 다른 지역일 수 있다는 주장을 뒷받침하는 것처럼 보였다. 타웅 아이의 인간과 유사한 특징은 어린 개체이기 때문에 나타나는 일시적인 현상이며, 성숙하면 사라질 것이라는 주장도 제기되었다. 그럼에도 불구하고 다트와 로버트 브룸은 ''Australopithecus''가 침팬지와는 거리가 멀고, 인간과 여러 가지 신체적, 행동적 유사성을 보인다고 주장하며 연구를 이어갔다.[8]
2. 2. 미세스 플레스와 추가 발견
로버트 브룸은 레이몬드 다트의 주장을 지지했다.[66] 1938년 브룸은 스테르크폰테인에서 발견된 뇌 용량이 485ml인 성인 두개골 화석(Sts 5)을 '플레시안트로푸스 트란스발렌시스'로 분류했다. 1947년 4월 17일, 브룸과 존 T. 로빈슨은 남아프리카 공화국의 스테르크폰테인 석회 채취장에서 중년 여성의 두개골 화석(Sts 5)을 발견했다.[67] 브룸은 이 화석 또한 '플레시안트로푸스 트란스발렌시스'로 분류했고, 언론에서는 이 화석을 "미세스 플레스"라고 불렀다. (하지만 이 화석은 현재 젊은 남성의 것으로 여겨지고 있다.) 이 뼈들에서도 레이몬드 다트가 보고한 것처럼 유인원의 특징이 보이지 않고, 오히려 인류와 가까운 특징이 보였기 때문에, 후에 오스트랄로피테쿠스 아프리카누스(A. africanus)로 재분류되었다.제2차 세계 대전 이후, 스테르크폰테인, 마카판스갓, 글래디스베일 등 남아프리카의 여러 지역에서 오스트랄로피테쿠스 아프리카누스 화석이 추가로 발견되면서 다트의 주장은 점차 설득력을 얻게 되었다. 1953년 필트다운 인 화석 조작 사건이 밝혀지면서, 오스트랄로피테쿠스 아프리카누스가 인류의 조상이라는 주장은 더욱 힘을 얻게 되었다.[8]
2. 3. 리틀 풋과 종 분류 논쟁
1994년, 남아프리카 스테르크폰테인 동굴에서 "리틀 풋"(StW 573)이라고 불리는 거의 완전한 오스트랄로피테쿠스 아프리카누스 골격 화석이 발견되었다.[37] 리틀 풋은 약 90%가 보존된, 지금까지 발견된 가장 완전한 초기 호미닌 골격이다.[37]리틀 풋 화석은 오스트랄로피테쿠스 아프리카누스의 해부학적 특징과 생활 방식에 대한 중요한 정보를 제공했다. 처음에는 330만 년 전으로 추정되었으나,[14] 2019년에는 367만 년 전으로 연대가 재측정되어 남아프리카에서 가장 오래된 오스트랄로피테쿠스 표본이 되었다.[15]
리틀 풋의 발견은 오스트랄로피테쿠스 아프리카누스 내 종 분화 논쟁을 다시 불러일으켰다. 1949년, 다트는 마카판스갓에서 발견된 여성 추정 얼굴 조각(MLD 2)을 "오스트랄로피테쿠스 프로메테우스(''A. prometheus'')"라는 새로운 종으로 분리할 것을 제안했다.[9] 그러나 1955년, 다트는 "''A. prometheus''"가 ''A. africanus''와 매우 유사하므로 동의어로 간주하는 데 동의했다.[11]
2008년, 고인류학자 로널드 J. 클라크는 StW 573(리틀 풋), StS 71 두개골, StW 505 두개골 등을 포함하여 "''A. prometheus''"를 다시 복원할 것을 제안했다.[13] 그러나 2018년, 고인류학자 리 로저스 버거와 존 D. 호크스는 "''A. prometheus''"가 ''A. africanus''와 구별되는 특징으로 적절하게 설명되지 않았다며 ''nomen nudum''("알려지지 않은 이름")으로 간주했다.[14]
3. 해부학적 특징
오스트랄로피테쿠스 아프리카누스는 오스트랄로피테쿠스 아파렌시스와 마찬가지로 두 발로 걸었지만, 팔이 다리보다 약간 더 길어 침팬지와 비슷한 특징을 보였다. 미세스 플레스와 "Sts 71" 두개골에서 볼 수 있듯이 머리 부분은 인간에 가깝고, 손가락은 굽기 쉬워 나무를 타는 데도 적합했다. 골반은 오스트랄로피테쿠스 아파렌시스보다 이족보행에 더 적합한 형태였다.[66]
찰스 다윈은 인류의 발상지가 아프리카라고 주장했지만, 20세기 전반에는 아시아 기원설이 주류였다.[68] 그러나 동아프리카의 Laetoli에서 사람과의 발자국 화석이 발견되면서, 메리 리키를 비롯한 리키 가문 과학자들은 아프리카 기원설을 지지했다.[69]
요추는 현생 인류와 마찬가지로 성적 이형성(남녀의 차이)을 보였으며, 여성의 골반은 임신 중에도 두 발로 걷는 데 적합했다.[70][71]
3. 1. 두개골 및 뇌

오스트랄로피테쿠스 아프리카누스(''A. africanus'')의 뇌 용량은 4개의 표본을 바탕으로 평균 정도였다. 이를 토대로 현대 영장류의 성체와 신생아 뇌 크기 추세를 이용하여 신생아 뇌 크기는 로 추정되었다. 이것이 정확하다면, ''A. africanus''는 총 뇌 크기의 약 38%로 태어났으며, 이는 30%인 인간보다 40%인 인간이 아닌 유인원과 더 유사하다는 것을 나타낸다.[25] 속귀는 인간처럼 달팽이관의 말단에서 헐거운 회전을 보이는 것뿐만 아니라 인간이 아닌 유인원과 같은 넓은 반고리관을 가지고 있다. 이러한 혼합은 속귀 해부학이 전정 기관(균형 감각)에 영향을 미치기 때문에 나무와 똑바로 걷는 습관적인 이동을 반영할 수 있다.[26]
''A. africanus''는 돌출된 턱을 가지고 있었고, 다소 움푹 들어간 얼굴(뺨이 부풀어 코가 움푹 들어간 부분의 아래에 위치함)과 뚜렷한 눈썹 융기를 가지고 있었다. 뇌두개골 양쪽으로 뻗어있는 측두선은 작은 볏으로 솟아있다. 송곳니는 인간이 아닌 유인원에 비해 크기가 줄어들었지만, 현대 인간의 송곳니보다 여전히 눈에 띄게 컸다. 다른 초기 호미닌과 마찬가지로, 어금니는 크고 두꺼운 법랑질을 특징으로 한다. 위턱에서 세 번째 어금니가 가장 큰 어금니이며, 아래턱에서는 두 번째 어금니이다. ''A. africanus''는 유인원과 같은 빠른 치아 발달 속도를 보였다.[8] ''A. africanus''는 두개골 특징에 대한 광범위한 변이를 가지고 있으며, 이는 일반적으로 수컷이 암컷보다 더 튼튼하다는 점에서 중간에서 높은 수준의 성적 이형성에 기인한다.[27]
3. 2. 골격 및 신체 비율
DIK-1-1의 ''오스트랄로피테쿠스 아파렌시스'' 골격을 바탕으로, 오스트랄로피테쿠스는 인간과 유사한 척추를 가지고 있었을 것으로 생각되며, 7개의 경추, 12개의 흉추, 그리고 (다른 초기 오스트랄로피테쿠스 골격을 바탕으로) 5개의 유연한 요추를 가지고 있었다.[32] StW 573의 목에 있는 환추는 머리를 돌리고 안정시키는 데 중요하며, 인간이 아닌 유인원과 더 유사하여 인간보다 위아래로 더 많이 움직일 수 있음을 나타낸다. 이러한 움직임은 오르기 쉬운 표면을 찾고 집중하는 수목 생활 종에게 중요하다. StW 573 환추는 침팬지 및 고릴라와 같이 어깨뼈를 움직이는 근육에 유사한 기계적 이점을 보이며, 이는 ''오스트랄로피테쿠스 아프리카누스''의 경추에서 척추 전만(척추의 정상적인 굽음)이 적었을 수 있음을 나타낼 수 있다. 그러나 이후의 StW 679는 인간의 환추와 유사한 점이 있어 유인원의 상태에서 점진적인 진화를 나타낼 수 있다.[33] StW 573은 ''오스트랄로피테쿠스 아파렌시스''와 인간과 달리 좁은 흉곽 입구를 가지고 있다. 쇄골은 비례적으로 매우 길며, 현대 인간의 쇄골과 절대 길이가 유사하다.[43]
현대 여성과 마찬가지로, 표본 StS 14에서는 L3–L5가 바깥쪽으로 굽어 있으며, 현대 남성과 같이 StW 431에서는 더 곧다. 이것은 임신 중에 똑바로 걷는 것을 돕기 위해 여성의 척추를 강화하는 것을 반영할 수 있다.[34] StS 14 부분 골격은 꽤 완전한 골반을 보존하고 있다. 루시 표본 (''오스트랄로피테쿠스 아파렌시스'')의 복원된 골반과 마찬가지로, 천골은 인간보다 상대적으로 평평하고 뒤쪽으로 더 기울어져 있으며, 골반강은 전반적으로 넓적한 모양을 하고 있었다. 이것은 신생아 머리 크기에 비해 넓은 산도를 나타낼 수 있으며, 따라서 회전이 없는 출산을 나타낼 수 있지만 (인간과 달리), 이에 대한 논쟁이 있다. 서 있을 때 천골과 요추 사이의 각도는 약 148.7°로 재구성되었으며, 이는 인간(118.3°)보다 침팬지(154.6°)와 훨씬 더 유사하다. 이는 ''오스트랄로피테쿠스 아프리카누스''의 서 있는 자세가 인간만큼 똑바르지 않았음을 나타낼 수 있다.[35]
''오스트랄로피테쿠스 아프리카누스''의 손과 팔은 모자이크 해부학적 특징을 보이며, 일부 측면은 인간과 더 유사하고 다른 측면은 비인간 유인원과 더 유사하다. 이것이 오스트랄로피테쿠스가 어느 정도 수목 생활을 했다는 의미인지, 아니면 이러한 특징이 단순히 인간-침팬지 마지막 공통 조상으로부터 물려받은 것인지는 불분명하다. 그럼에도 불구하고, ''오스트랄로피테쿠스 아프리카누스''는 ''오스트랄로피테쿠스 아파렌시스''보다 더 유인원과 같은 상지 해부학적 특징을 보이며, 어느 정도는 수목 생활을 했을 것으로 해석된다. 수목 생활 영장류처럼 손가락은 굽어 있고, 팔은 비교적 길며, 어깨는 으쓱하는 위치에 있다. ''오스트랄로피테쿠스 아프리카누스''의 어깨는 오랑우탄의 어깨와 가장 유사하며, 머리 위로 들어 올려 무게를 지탱하고 유지하는 데 적합하다.[36] 그러나 StW 573의 오른쪽 쇄골은 인간과 같이 뚜렷한 S자형(구불구불한) 곡선을 가지고 있어, 상완골에 대한 견갑골을 안정시키는 인간과 유사한 모멘트 팔을 나타낸다.[43] ''오스트랄로피테쿠스 아프리카누스''의 팔뼈는 등반에 유용한 강력한 근육과 일치한다. 그럼에도 불구하고, 상완 지수(전완 대 상완골 비율)는 82.8–86.2(침팬지와 인간의 중간)로, 더 오래된 호미닌 ''아르디피테쿠스 라미두스''에서 전완 길이의 감소를 나타낸다.[37] 엄지와 손목은 정밀 파지와 엄지와 손가락 사이의 강력한 대립을 통해 인간과 유사한 기능을 나타낸다. 이러한 파지의 채택은 일반적으로 효율적인 등반과 수목 생활을 희생하고 도구 제작에 적응한 것으로 해석된다.[38]

다리뼈는 ''오스트랄로피테쿠스 아프리카누스''가 습관적으로 두 발로 걷는다는 것을 분명히 보여주지만, 경골의 일부 측면은 유인원과 유사하며, 이는 다리 근육이 인간 상태로 완전히 재구성되지 않았음을 나타낼 수 있다. 이것이 맞다면, 그 기능적 의미는 불분명하다.[37] 고관절의 섬유주 골은 뚜렷하게 인간과 유사하며, 이는 장기간의 수목 활동에 필요한 큰 정도의 고관절 부하와 일치하지 않는다.[39] 경골은 인간과 유사한 각도로 발과 만나며, 이는 습관적인 두 발 보행에 필요하다. 결과적으로 발목은 비인간 유인원만큼 등반 활동에 능숙하지 않았다.[40] 그러나 현대 콩고 트와 수렵 채집인은 특정 골격 적응 대신 위장근(종아리)의 더 긴 섬유로 인해 나무를 오르는 동안 침팬지와 유사한 각도를 얻을 수 있다.[41] 발목뼈의 일부 측면은 유인원과 유사하여 걷는 효율성에 영향을 미쳤을 수 있다. ''오스트랄로피테쿠스 아프리카누스''의 발 요소는 주로 Sterkfontein Member 4의 유해에서 알려져 있다. 발은 발 중앙부가 뻣뻣하고 중간 발목 골절이 없다는 점에서 인간과 유사하다(이것은 비인간 유인원이 발의 나머지 부분에서 독립적으로 발꿈치를 들어 올릴 수 있게 해준다). ''오스트랄로피테쿠스 아프리카누스''는 인간과 같이 내전된 큰 발가락(손재주가 없음)을 가지고 있었지만, 발의 옆면을 사용하여 큰 발가락으로 밀지 않았을 가능성이 있다. StW 573은 내전된 큰 발가락을 가진 가장 오래된 호미닌 표본이다. 표본 StW 355는 알려진 모든 호미닌 중 가장 굽은 근위 발 지골이며, 오랑우탄 및 시앙과 더 유사하다.[42]
StW 573의 팔 길이는 약 53.4cm, 다리 길이는 61.5cm였다. 즉, 팔 길이는 다리 길이의 86.9%였다. 그녀는 팔이 다리와 거의 같은 길이임을 확실하게 보여주는 최초이자 유일한 초기 호미닌 표본이다. 그럼에도 불구하고, 이러한 비율은 비인간 유인원보다 인간과 더 유사하며, 인간은 64.5–78%, 침팬지는 약 100%, 고릴라는 100–125%, 오랑우탄은 135–150.9%이다.[43]
요추가 현생 인류와 마찬가지로 성적 이형성(남녀의 차이)을 가지고 있었다는 것이 알려진 최초의 호미니드는 *오스트랄로피테쿠스 아프리카누스*이다. 이 골반은 여성이 임신 중에 두 발로 걷는 데 적합하다.[70][71]
4. 고생물학
오스트랄로피테쿠스 아프리카누스는 다재다능한 잡식성으로, 매우 다양한 음식을 섭취했다. 탄소 동위원소 분석 결과, 이들은 풀, 씨앗, 근경, 지하 저장 기관(덩이줄기 등) 또는 풀을 먹는 무척추동물(예: 메뚜기 또는 흰개미), 방목하는 포유류, 곤충을 먹는 동물 등 상당량의 C4 사바나 식물을 섭취했다.[46][44] 대부분의 영장류는 C4 식물을 먹지 않는다. 얼굴 해부학적 구조는 치아로 작은 견과류와 씨앗 같은 단단한 물체를 부수거나, 한 번에 많은 양의 음식을 처리하는 데 유리하도록 앞어금니에 높은 스트레스를 가하는 데 적응한 것으로 보인다. 그러나 어금니의 미세 마모 분석 결과, 작고 단단한 음식은 드물게 먹었으며, 이는 먹이가 부족한 시기에 비상식으로 먹었을 가능성이 있다.[45] 일부 화석에서는 감귤류와 같은 산성 음식을 씹은 흔적이 발견되기도 했다.[49]
2011년 스트론튬 동위원소 연구에서 남아프리카 스테르크폰테인 계곡에서 발견된 ''A. africanus''의 치아를 분석했다. 그 결과, 작은 치아는 암컷을, 큰 치아는 수컷을 나타낸다고 가정했을 때, 암컷이 태어난 곳을 떠날 가능성이 더 높았다(부계 거주).[51] 이는 현대 사람족의 분산 패턴과 유사하며, 고릴라 및 다른 영장류의 하렘 사회와는 달리 여러 수컷을 기반으로 한 친족 사회를 이루었을 가능성을 시사한다. 수컷의 작은 송곳니는 암컷의 송곳니에 비해 비인간 호미닌보다 수컷 간의 공격성이 훨씬 낮았음을 시사한다. 수컷은 계곡에서 멀리 벗어나지 않은 것으로 보이는데, 이는 작은 생활권이 있었거나, 동굴의 풍부함이나 식물 성장에 관련된 요인으로 인해 백운암 지형을 선호했음을 나타낼 수 있다.[51]
10개의 ''오스트랄로피테쿠스 아프리카누스'' 표본 중 7개에서 치주 질환(잇몸 질환으로 인해 치아를 지지하는 뼈가 닳아 없어지는 현상)이 경미하거나 중간 정도로 나타났다.[52] 어린 개체인 STS 24a는 입 오른쪽에서 극심한 치주 질환이 진단되었으며, 이는 감염 부위 주변의 병적인 뼈 성장과 주기적인 세균 감염 및 그로 인한 염증 기간 동안 첫 번째와 두 번째 오른쪽 어금니의 이동을 유발했다. 이 개체는 턱의 왼쪽을 사용하여 씹는 것을 선호했던 것으로 보인다. 치주 질환은 특히 생애 마지막 해에 씹는 것을 심각하게 방해했을 것이며, 이 개체는 생존하기 위해 오랫동안 무리 구성원에게 의존했을 가능성이 있다.[53] 1992년, 인류학자 제프리 레이몬드 피스크와 가브리엘레 마초는 왼쪽 발목뼈 Stw 363에서 뒤꿈치뼈(보존되지 않음)의 치유된 종골 골절의 증거를 발견했는데, 그들은 이것이 나무에서 떨어져서 발생했다고 믿었다. 만약 이것이 사실이라면, 이 개체는 왼쪽 다리의 기능을 많이 잃었음에도 불구하고 오랫동안 생존할 수 있었다. 그러나 그들은 또한 화석화 과정에서 방해석 침전과 결정화에 의해 유사한 손상이 발생했을 가능성도 지적했다. 종골 골절은 인간에게서 기록되었으며, 수목성 영장류에게서 매우 자주 나타난다.[54]
4. 1. 식생
오스트랄로피테쿠스 아프리카누스는 다재다능한 잡식성으로, 매우 다양한 음식을 섭취했다. 탄소 동위원소 분석 결과, 이들은 풀, 씨앗, 근경, 지하 저장 기관(덩이줄기 등) 또는 풀을 먹는 무척추동물(예: 메뚜기 또는 흰개미), 방목하는 포유류, 곤충을 먹는 동물 등 상당량의 C4 사바나 식물을 섭취했다.[46][44] 대부분의 영장류는 C4 식물을 먹지 않는다.얼굴 해부학적 구조는 치아로 작은 견과류와 씨앗 같은 단단한 물체를 부수거나, 한 번에 많은 양의 음식을 처리하는 데 유리하도록 앞어금니에 높은 스트레스를 가하는 데 적응한 것으로 보인다. 그러나 어금니의 미세 마모 분석 결과, 작고 단단한 음식은 드물게 먹었으며, 이는 먹이가 부족한 시기에 비상식으로 먹었을 가능성이 있다.[45]
일부 화석에서는 감귤류와 같은 산성 음식을 씹은 흔적이 발견되기도 했다.[49]
4. 2. 사회
2011년 스트론튬 동위원소 연구에서 백운암으로 이루어진 스테르크폰테인 계곡에서 발견된 ''A. africanus''의 치아를 분석했다. 그 결과, 작은 치아는 암컷을, 큰 치아는 수컷을 나타낸다고 가정했을 때, 암컷이 태어난 곳을 떠날 가능성이 더 높았다(부계 거주).[51] 이는 현대 사람족의 분산 패턴과 유사하며, 고릴라 및 다른 영장류의 하렘 사회와는 달리 여러 수컷을 기반으로 한 친족 사회를 이루었을 가능성을 시사한다. 그러나 수컷의 작은 송곳니는 암컷의 송곳니에 비해 비인간 호미닌보다 수컷 간의 공격성이 훨씬 낮았음을 시사한다. 수컷은 계곡에서 멀리 벗어나지 않은 것으로 보이는데, 이는 작은 생활권이 있었거나, 동굴의 풍부함이나 식물 성장에 관련된 요인으로 인해 백운암 지형을 선호했음을 나타낼 수 있다.[51]4. 3. 병리
10개의 ''오스트랄로피테쿠스 아프리카누스'' 표본 중 7개에서 치주 질환(잇몸 질환으로 인해 치아를 지지하는 뼈가 닳아 없어지는 현상)이 경미하거나 중간 정도로 나타났다.[52] 어린 개체인 STS 24a는 입 오른쪽에서 극심한 치주 질환이 진단되었으며, 이는 감염 부위 주변의 병적인 뼈 성장과 주기적인 세균 감염 및 그로 인한 염증 기간 동안 첫 번째와 두 번째 오른쪽 어금니의 이동을 유발했다. 마찬가지로, 이 개체는 턱의 왼쪽을 사용하여 씹는 것을 선호했던 것으로 보인다. 치주 질환은 특히 생애 마지막 해에 씹는 것을 심각하게 방해했을 것이며, 이 개체는 생존하기 위해 오랫동안 무리 구성원에게 의존했을 가능성이 있다.[53]1992년, 인류학자 제프리 레이몬드 피스크와 가브리엘레 마초는 왼쪽 발목뼈 Stw 363에서 뒤꿈치뼈(보존되지 않음)의 치유된 종골 골절의 증거를 발견했는데, 그들은 이것이 나무에서 떨어져서 발생했다고 믿었다. 만약 이것이 사실이라면, 이 개체는 왼쪽 다리의 기능을 많이 잃었음에도 불구하고 오랫동안 생존할 수 있었다. 그러나 그들은 또한 화석화 과정에서 방해석 침전과 결정화에 의해 유사한 손상이 발생했을 가능성도 지적했다. 종골 골절은 인간에게서 기록되었으며, 수목성 영장류에게서 매우 자주 나타난다.[54]
5. 고환경
남아프리카의 오스트랄로피테쿠스 아프리카누스는 다양한 서식지가 있는 지역에 살았던 것으로 보인다. 스테르크폰테인에서 발견된 덩굴 화석과 야생 동물 군집은 관목 사바나, 열린 삼림지, 초원 등 다양한 환경을 나타낸다. 관목 ''Anastrabe integerrima'' 화석은 인류의 요람 지역이 플리오-플라이스토세에 더 많은 강우량을 받았음을 시사하며, 초원으로 둘러싸인 갤러리 숲의 특징을 가졌을 수 있음을 보여준다.[55] 타웅 역시 습하고 폐쇄된 환경이었던 것으로 보인다.[56]
오스트랄로피테신과 초기 ''호모''는 침팬지처럼 일평균 기온이 25°C이고 밤에는 10°C로 떨어지는, 더 시원한 환경을 선호했을 가능성이 있다.[57]
약 207만 년 전, ''P. robustus''와 ''H. erectus''가 출현하기 직전에 ''A. africanus''는 인류의 요람에서 멸종되었다. 남아프리카는 주요 기후 변동성과 변동의 시작 전까지, 그리고 아마도 ''호모''와 ''파란트로푸스''와의 경쟁 속에서 ''오스트랄로피테쿠스''의 피난처였을 수 있다.[62]
5. 1. 서식지
남아프리카의 오스트랄로피테쿠스 아프리카누스는 다양한 서식지가 있는 지역에 살았던 것으로 보인다. 스테르크폰테인에서 발견된 덩굴 화석과 야생 동물 군집은 관목 사바나, 열린 삼림지, 초원 등 다양한 환경을 나타낸다. 관목 ''Anastrabe integerrima'' 화석은 인류의 요람 지역이 플리오-플라이스토세에 더 많은 강우량을 받았음을 시사하며, 초원으로 둘러싸인 갤러리 숲의 특징을 가졌을 수 있음을 보여준다.[55] 타웅 역시 습하고 폐쇄된 환경이었던 것으로 보인다.[56]오스트랄로피테신과 초기 ''호모''는 침팬지처럼 일평균 기온이 25°C이고 밤에는 10°C로 떨어지는, 더 시원한 환경을 선호했을 가능성이 있다.[57]
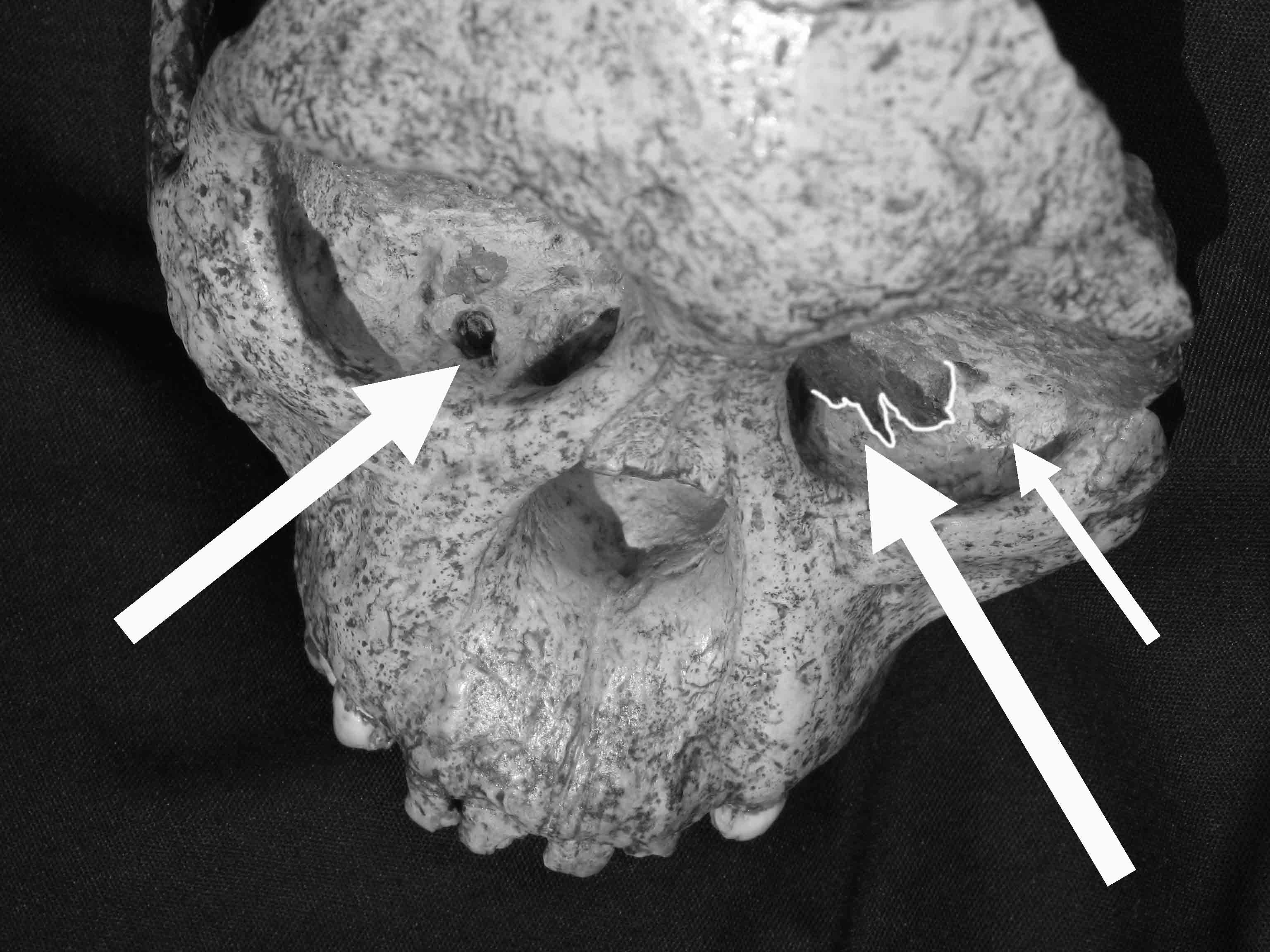
1983년 남아프리카 고생물학자 찰스 킴벌린 브레인은 오스트랄로피테신의 뼈가 대형 육식 동물의 활동으로 동굴에 축적되었다는 가설을 제시했다. 스테르크폰테인 멤버 4에서 발견된 ''A. africanus'' 화석은 고양이과 동물에 의해 축적되었을 가능성이 높지만, 사냥 하이에나와 자칼도 역할을 했을 수 있다.[59] 타웅 아이의 두개골에는 현대 관머리수리가 입힌 것과 유사한 손상이 발견되었는데, 이는 맹금류에 의해 살해되었음을 나타낸다.[60][61]
5. 2. 포식자
타웅 아이의 두개골에는 맹금류에게 공격당해 안와에 손상을 입은 흔적이 발견되었다.[60][61] 남아프리카 고생물학자 찰스 킴벌린 브레인은 오스트랄로피테신 뼈가 대형 육식 동물의 활동으로 동굴에 축적되었다는 가설을 세웠다.[58] 스테르크폰테인 멤버 4에서 발견된 ''A. africanus'' 화석은 고양이과 동물에게 사냥 당했을 가능성이 높지만, 사냥 하이에나와 자칼도 사냥에 관여했을 수 있다.[59]
6. 진화 계통 및 멸종
찰스 다윈은 1871년 ''인간의 유래''에서 인류가 아프리카에서 진화했다고 주장했으며, ''A. 아프리카누스''는 이를 뒷받침하는 최초의 증거였다.[8] 초기에는 고릴라나 침팬지 계통으로 여겨지기도 했으나, 제2차 세계 대전 이후 여러 연구와 발견을 통해 ''A. africanus''가 인간 조상으로 분류되어야 한다는 견해가 널리 받아들여졌다. 1947년에는 가장 완전한 두개골인 STS 5("Mrs. Ples")가 발견되었고,[7] 1953년에는 필트다운 인이 조작된 것임이 밝혀지면서 ''A. africanus''의 인류 조상설은 더욱 힘을 얻었다.[8]
''A. africanus''와 다른 호미닌과의 관계는 불분명하다.[20] 20세기 후반 아프리카에서 초기 홍적세 ''호모''가 발견되면서 ''A. africanus''는 ''호모''의 조상으로 여겨졌다. 이후 발견된 ''A. afarensis''는 ''A. africanus''는 ''P. robustus''의 조상, ''A. afarensis''는 ''호모''와 ''A. africanus''/''P. robustus''의 공통 조상이라는 가설이 제시되기도 하였다.[21]
약 207만 년 전, 파란트로푸스 로부스투스와 호모 에렉투스가 출현하기 직전에 남아프리카에서 오스트랄로피테쿠스 아프리카누스는 멸종했다. 남아프리카는 주요 기후 변동과 변동이 시작되기 전까지 오스트랄로피테쿠스의 피난처였을 수 있으며, 아마도 호모 속(屬)과 파란트로푸스 속(屬)과의 경쟁에서 밀려났을 것으로 추정된다.[62]
6. 1. 다른 종과의 관계
찰스 다윈은 1871년 저서 ''인간의 유래''에서 인류가 아프리카에서 진화했다고 주장했으며, ''A. 아프리카누스''는 이를 뒷받침하는 최초의 증거였다.[8] 그러나 당시의 일반적인 학설은 큰 뇌와 인간과 같은 특징이 이족 보행보다 먼저 진화했다는 것이었다. 따라서 ''A. africanus''는 고릴라나 침팬지 계통으로 여겨졌으며, 특히 아서 키스 경이 이러한 주장을 펼쳤다.[8]찰스 도슨의 1912년 사기극인 필트다운 인 발견과 중국에서의 베이징 인(''Homo erectus pekinensis'') 발견은 인류 기원이 아프리카가 아니라는 주장에 힘을 실었다. 타웅 아이의 인간과 같은 특징은 어린 개체의 특징이며 성숙하면 사라질 것이라고 여겨졌다. 그럼에도 로버트 브룸 등은 ''Australopithecus''가 침팬지와 거리가 멀고 인간과 유사한 신체적, 행동적 특징을 보인다고 주장했다.[8]
제2차 세계 대전 이후, 로버트 브룸과 Gerrit Willem Hendrik Schepers의 연구, 윌프리드 르 그로스 클라크 경의 논문 등을 통해 ''A. africanus''가 인간 조상으로 분류되어야 한다는 견해가 널리 받아들여졌다.[8] 1947년에는 가장 완전한 두개골인 STS 5("Mrs. Ples")가 발견되었고,[7] 1953년에는 필트다운 인이 조작된 것임이 밝혀지면서 ''A. africanus''의 인류 조상설은 더욱 힘을 얻었다.[8]
현재 오스트랄로피테쿠스 속의 분류는 명확하지 않다. ''Australopithecus''는 계통 분류군으로, 구성원 간의 관계보다는 유사한 생리학적 특징으로 묶인다.[20] ''A. africanus''와 다른 호미닌과의 관계는 불분명하다.[20] 20세기 후반 아프리카에서 초기 홍적세 ''호모''가 발견되면서 ''A. africanus''는 ''호모''의 조상으로 여겨졌다. 1978년 발견된 ''A. afarensis''는 당시 가장 오래된 호미닌으로, ''A. africanus''는 ''P. robustus''의 조상, ''A. afarensis''는 ''호모''와 ''A. africanus''/''P. robustus''의 공통 조상이라는 가설이 제시되었다.[21] ''A. africanus''가 동아프리카의 ''Paranthropus'' 종이 아닌 ''P. robustus''와 가깝거나,[22] 모든 ''Paranthropus''의 조상이라는 가설도 있다.[23] 인류의 요람에서 발견된 ''A. sediba''는 ''A. africanus''의 조상이자 ''호모''의 조상으로 추정되어, ''A. africanus''가 ''호모''의 조상일 가능성을 시사한다.[24]
''A. afarensis''처럼 A. 아프리카누스는 이족 보행을 했고 팔이 다리보다 약간 길었다. 이는 침팬지와 유사하다. "Mrs. Ples"와 "Sts 71" 두개골에서 볼 수 있듯 머리 부분은 인간과 유사하며, 손가락은 굽혀져 나무 타기에 적합했다.
파란트로푸스속이 A. 아프리카누스에서 진화했다는 주장도 있다. 파란트로푸스 로부스투스는 고릴라와 유사한 두개골을 가져 A. 아프리카누스보다 강한 씹는 힘을 가졌을 것으로 추정된다. 반면 A. 아프리카누스의 두개골은 침팬지와 유사하다. 뇌 용량은 400-500ml로 유인원 수준의 지능을 가졌을 것으로 보인다.[66] A. 아프리카누스의 골반은 오스트랄로피테쿠스 아파렌시스보다 이족보행에 더 적합했다.
6. 2. 멸종
약 207만 년 전, 파란트로푸스 로부스투스와 호모 에렉투스가 출현하기 직전에 남아프리카에서 오스트랄로피테쿠스 아프리카누스는 멸종했다. 남아프리카는 주요 기후 변동과 변동이 시작되기 전까지 오스트랄로피테쿠스의 피난처였을 수 있으며, 아마도 호모 속(屬)과 파란트로푸스 속(屬)과의 경쟁에서 밀려났을 것으로 추정된다.[62]참조
[1]
웹사이트
Australopithecus africanus
https://humanorigins[...]
Smithsonian Institution
2024-01-03
[2]
논문
The Hypothesized Osteodontokeratic Culture of the Australopithecinae: A Look at the Evidence and the Opinions
[3]
논문
The Predatory Transition from Ape to Man
http://www.users.mia[...]
[4]
간행물
New Fossil Anthropoid Skull from South Africa
Nature
[5]
간행물
On some new Pleistocene mammals from limestone caves of the Transvaal
South African Journal of Science
[6]
논문
The Pleistocene Anthropoid Apes of South Africa
[7]
논문
Further Remains of the Sterkfonstein Ape-Man, ''Plesianthropus''
[8]
논문
Ape-Like ''Australopithecus'' After Seventy Years: Was It a Hominid?
[9]
논문
The cranio-facial fragments of ''Australopithecus prometheus''
[10]
논문
The Second, or Adult, Female Mandible of ''Australopithecus prometheus''
[11]
논문
''Australopithecus prometheus'' and ''Telanthropus capensis''
[12]
논문
A New Species of the Genus ''Australopithecus'' (Primates: Hominidae) from the Pliocene of Eastern Africa
https://www.biodiver[...]
1978
[13]
논문
Latest information on Sterkfontein's ''Australopithecus'' skeleton and a new look at ''Australopithecus''
http://www.scielo.or[...]
[14]
논문
''Australopithecus prometheus'' is a ''nomen nudum''
[15]
논문
The skull of StW 573, a 3.67 Ma ''Australopithecus prometheus'' skeleton from Sterkfontein Caves, South Africa
[16]
논문
Gladysvale: First early hominid site discovered in South Africa since 1948
[17]
논문
Cosmogenic nuclide dating of Australopithecus at Sterkfontein, South Africa
2022-07-05
[18]
논문
The Long-Term Conservation of the ''Australopithecus''-bearing Member 4 Excavation Walls at the Sterkfontein Caves, South Africa
[19]
논문
The elusive 'second species' in Sterkfontein Member 4: The dental metrical evidence
https://www.research[...]
[20]
논문
Hominin Taxonomy and Phylogeny: What's In A Name?
https://www.nature.c[...]
[21]
논문
A Systematic Assessment of Early African Hominids
[22]
논문
''Paranthropus boisei'': Fifty years of evidence and analysis
https://mds.marshall[...]
[23]
논문
Multiplying genera versus moving species: a new taxonomic proposal for the family Hominidae
[24]
논문
Characterizing the Evolutionary Path(s) to Early Homo
[25]
논문
Brain size at birth throughout human evolution: A new method for estimating neonatal brain size in hominins
[26]
논문
The bony labyrinth of StW 573 ("Little Foot"): Implications for early hominin evolution and paleobiology
[27]
논문
Sexual dimorphism in the face of ''Australopithecus africanus''
[28]
논문
Body Size and Proportions in Early Hominids
[29]
논문
The evolution of body size and shape in the human career
[30]
논문
Femoral Lengths and Stature in Plio-Pleistocene Hominids
[31]
논문
Long-term patterns of body mass and stature evolution within the hominin lineage
[32]
논문
Thoracic Vertebral Count and Thoracolumbar Transition in ''Australopithecus afarensis''
[33]
논문
The atlas of StW 573 and the late emergence of human-like head mobility and brain metabolism
[34]
논문
Fetal load and the evolution of lumbar lordosis in bipedal hominins
http://nrs.harvard.e[...]
[35]
논문
A new reconstruction of Sts 14 pelvis (''Australopithecus africanus'') from computed tomography and three-dimensional modeling techniques
[36]
논문
Like Father, Like Son: Assessment of the Morphological Affinities of A.L. 288–1 (''A. afarensis''), Sts 7 (''A. africanus'') and Omo 119–73–2718 (''Australopithecus'' sp.) through a Three-Dimensional Shape Analysis of the Shoulder Joint
[37]
논문
The Long Limb Bones of the StW 573 ''Australopithecus'' Skeleton from Sterkfontein Member 2: Descriptions and Proportions
[38]
논문
Human-like hand use in ''Australopithecus africanus''
[39]
논문
Evidence for habitual climbing in a Pleistocene hominin in South Africa
[40]
논문
Trabecular Evidence for a Human-Like Gait in ''Australopithecus africanus''
[41]
논문
Tree climbing and human evolution
[42]
논문
One small step: A review of Plio-Pleistocene hominin foot evolution
[43]
논문
Functional Anatomy, Biomechanical Performance Capabilities and Potential Niche of StW 573: an ''Australopithecus'' Skeleton (circa 3.67 Ma) From Sterkfontein Member 2, and its significance for The Last Common Ancestor of the African Apes and for Hominin Origin
[44]
논문
The carbon isotope ecology and diet of ''Australopithecus africanus'' at Sterkfontein, South Africa
[45]
논문
The feeding biomechanics and dietary ecology of ''Australopithecus africanus''
[46]
논문
Isotopic Evidence for the Diet of an Early Hominid, ''Australopithecus africanus''
[47]
논문
Root caries on a ''Paranthropus robustus'' third molar from Drimolen
https://www.biorxiv.[...]
[48]
논문
Dental caries in human evolution: frequency of carious lesions in South African fossil hominins
[49]
논문
Root grooves on two adjacent anterior teeth of ''Australopithecus africanus''
http://researchonlin[...]
[50]
논문
Elemental signatures of ''Australopithecus africanus'' teeth reveal seasonal dietary stress
[51]
논문
Strontium isotope evidence for landscape use by early hominins
[52]
논문
The Hard Evidence of Alveolar Bone Loss in Early Hominids of Southern Africa
[53]
논문
Paleopathology in ''Australopithecus africanus'': A suggested case of a 3-million-year-old prepubertal periodontitis
[54]
논문
Evidence of a healed compression fracture in a Plio-Pleistocene hominid talus from Sterkfontein, South Africa
[55]
논문
Pliocene Fossil Woods from an Early Hominid Cave Deposit, Sterkfontein, South Africa
https://www.research[...]
[56]
논문
Reconstruction the Paleoecology of Taung, South Africa from Low Magnification of Dental Microwear Features in Fossil Primates
[57]
논문
Bipedality and hair loss in human evolution revisited: The impact of altitude and activity scheduling
[58]
서적
The Hunters Or the Hunted?: An Introduction to African Cave Taphonomy
https://books.google[...]
University of Chicago Press
[59]
논문
An ecological reassessment of the southern African carnivore guild: a case study from Member 4, Sterkfontein, South Africa
[60]
논문
Brief Communication: Predatory bird damage to the Taung type-skull of ''Australopithecus africanus'' Dart 1925
[61]
논문
Further evidence for eagle predation of, and feeding damage on, the Taung child
http://www.scielo.or[...]
[62]
논문
Contemporaneity of ''Australopithecus'', ''Paranthropus'', and early ''Homo erectus'' in South Africa
[63]
웹사이트
Human Ancestors Hall: Tree
http://www.mnh.si.ed[...]
[64]
웹사이트
Australopithecus africanus
http://www.archaeolo[...]
[65]
웹사이트
Raymond Dart and our African origins
http://www.press.uch[...]
[66]
웹사이트
Primate Origins
http://www.anth.ucsb[...]
[67]
웹사이트
John T. Robinson
http://archaeology.a[...]
[68]
웹사이트
New Ideas About Human Migration From Asia To Americas
http://www.scienceda[...]
[69]
웹사이트
Apologetics Press - Human Evolution and the “Record of the Rocks”
http://www.apologeti[...]
[70]
뉴스
A pregnant woman's spine is her flexible friend
http://news.independ[...]
The Independent
2007-12-13
[71]
뉴스
Why Pregnant Women Don't Tip Over.
http://news.national[...]
National Geographic News
2007-12-12
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com